Autonomy and Adaptive Behaviour
Towards a naturalized and biologically
inspired definition of behavioural adaptive autonomy
Xabier Barandiaran
xabier @ barandiaran.net
http://barandiaran.net
Autonomía Situada
grey-walter @ sindominio.net
http://sindominio.net/autonomiasituada
08-07-03
Within a naturalized dynamical approach to adaptive behaviour and
cognition, behavioural adaptive autonomy is defined as:
homeostatic maintenance of essential variables under viability
constraints through self-modulating behavioural coupling with the
environment, hierarchically decoupled from metabolic (constructive)
processes. This definition allows for a naturalized notion of
normative functionality, structurally and interactivelly emergent.
We argue that artificial life techniques such as evolutionary
simulation modelling provide a workable methodological framework for
philosophical research on complex adaptive behaviour.
Autonomy, adaptive behaviour, philosophy of biology, philosophy of
cognitive science, cognition, dynamical systems, naturalization.
Autonomy and Adaptive Behaviour. Towards a naturalized and biologically
inspired definition of behavioural adaptive autonomy. v.0.8
Copyright © 2003 Xabier Barandiaran.
Copyleft 2003 Xabier Barandiaran:
The licensor permits others to copy, distribute,
display, and perform the work. In return, licensees must give the
original author credit. The licensor permits others to copy,
distribute, display, and perform the work. In return, licensees may
not use the work for commercial purposes --unless they get the
licensor's permission. The licensor permits others to
distribute derivative works only under a license identical to the
one that governs the licensor's work.
This is not mean to be the full license but a short guide of it.
The full license can be found at:
http://creativecommons.org/licenses/by-nc-sa/1.0/legalcode
Xabier Barandiaran (2003) Autonomy and Adaptive Behaviour. Towards a naturalized and biologically
inspired definition of behavioural adaptive autonomy. v.0.8. url:
http://barandiaran.net/textos/auto/auto.pdf
In this paper we will try:
- to explain the significance of autonomy (dynamically
considered) for adaptive behaviour and cognitive science (section 2),
- to clarify the different aspects of autonomy in
relation to adaptive behaviour (section 3),
- to specify a notion of behavioural adaptive autonomy
and normative functionality in the framework of a dynamical
approach to cognition and adaptive behaviour (section 4),
- to justify (evolutionary) simulation modelling as a workable
methodological framework for the study of behavioural adaptive
autonomy (section 5) and
- to extract some consequences of behavioural adaptive
autonomy for a naturalized definition of cognition while
illustrating some discusion with recent work on evolutionary
simulation modelling (section 6).
The main thesis is:
- that adaptive behavioural autonomy shall
be defined as:
homeostatic maintenance of essential variables under
viability constraints through self-modulating behavioural coupling
with the environment, hierarchically decoupled from metabolic
(constructive) processes
- and that this definition of
behavioural adaptive autonomy should serve as a lower boundary for a
naturalist characterization of cognition as emergent from life but
distinct from it.
In addition we provide a set of formal definitions at the end of the
paper.
The term autonomy and autonomous has been
largely used in cognitive science and robotics (Maes, 1991)
to describe an agent embodied and situated in the `real world' and
without external energy supply; we consider
that a deeper sense of autonomy (as self-maintenance)
allows for a richer characterization of cognition and
adaptive behaviour. In this line the work of Varela (1979)
provides a deeper sense of autonomy, recently developed by
Ruiz-Mirazo and Moreno (2000) in the dimension of basic
(autopoietic) autonomy and by Christensen and Hooker (2002) in the
dimension of adaptation and cognition conceptualized through the
notion of self-directedness; while Bickhard (2000) has analysed
the consequences of autonomy for functional and representational
normativity1. FALTA ENRIQUECER ESTE PÁRRAFO CON DIFERENTES
AUTORES Y UNA BREVE HISTORIA DEL USO DEL CONCEPTO DE AUTONOMÍA EN LA
SIMULACIÓN DE CONDUCTA ADAPTATIVA.
But the relation between basic (autopoietic or self-maintaining)
autonomy and behavioural or cognitive autonomy deserves some
clarification in order to be introduced in dynamical system theory
(as a conceptual framework) and evolutionary simulation modelling of
adaptive behaviour (as a workable methodological framework). This is
the main goal of this paper. At the same time by conceptualizing and
modelling autonomy within the dynamical approach to cognitive science
(van Gelder, 1998) and adaptive behaviour (Beer, 1997) a number of
important goals could be achieved:
- To provide a autonomous normative criteria to interpret
and evaluate adaptive and cognitive functionality,
solving the frame of reference problem
(Clancey, 1989) of computational functionalist
approaches (Block, 1996).
- To naturalize such normative criteria
on the dynamical organization of neural and
interactive processes (and their relation with self-maintenance)
giving rise to adaptive/cognitive behaviour
as proposed by Bickhard, Christensen and Hooker
(Christensen and Hooker, 2002; Cristensen and Hooker, 1999; Christensen and Bickhard, 2002);
without
recursion to evolutionary functionalism
(Millikan, 1989a,b)
or an absolute external observer in order to attribute
structural/functional relations within the organism and
between the organism and its environment.
- To integrate mechanistic, embodied and
situated (interactive) explanations without recursion to
pre-specified functional/behavioural primitives, thus integrating
behavioural and structural complexity in a workable
methodological framework. This will satisfy
holistic, organismic or organizational criticisms
(Gilbert and Sarkar, 2000) to traditional functionalist
perspectives on cognition while providing synthetic and analytic
criteria for advances in scientific research (thus avoiding the
often questioned solipsist danger of such approaches).
The origin of the word autonomy comes from the Greek
auto-nomos (self-law). We can thus provide an intuitive first
notion of autonomous systems as those producing their own
laws2.
But this notion requires a previous notion of self: autonomous
systems must first produce their own identity; i.e. autonomous
systems are primarily those whose basic organization is that of a
self-sustaining, self-constructing entity over time and space.
Basic autonomy (Ruiz-Mirazo and Moreno, 2000) is the organization by
which far from equilibrium and thermodynamically open systems
adaptively generate internal and interactive constraints to modulate
the
flow of matter and energy required for their self-maintenance. Two
equally fundamental but distinct aspects of basic autonomy can be
distinguished:
- a) constructive:
- generation of internal
constraints to control the internal flow of matter and energy for
self-maintenance. In this sense the autonomous system can be
understood as a highly recursive network of processes that produces
the components that constitute the network itself
(Maturana and Varela, 1980). Metabolism is the main expression of
this constructive aspect.
- b) interactive:
- the generation of
interactive constraints modulating the boundary conditions
of the system to assure the necessary flow of energy and matter
between the systems and its environment for self
maintenance (unlike dissipative structures which hold their
organization only under a restricted set of external conditions that
the system cannot modify). The membrane of a cell, controll of
behaviour or breathing are characteristic examples of this
interactive constraint generation.
On this basis we can define constructive closure as the
satisfaction of constructive constraint generation and
interactive closure as the satisfaction of interactive
constraint generation for self maintenance.
It is the satisfaction of closure conditions
that defines the function (Collier, 1999) of internal and
interactive processes. Functionality is, thus, picked
up at the level of their contribution to self-maintenance and
not, as evolutionary functionalism proposes, at the level
of selective history; nor, as computational functionalists defended,
as externally (heteronomously) interpreted causal relations between
computational states (and, when cognition is involver,
their representational relation with
external ``states of affairs''). For, of course,
contribution to self-maintenance is evolutionarily advantageous; but
autonomy is to be seen not as a pure outcome of evolutionary
processes but as the condition of possibility of such process.
Functions become normative by means of the dynamic
presupossition of that process in the overall organization of the
system (Bickhard, 2000; Christensen and Bickhard, 2002). In other
words because constructive and interactive functional processes are
the condition of possibility of autonomous systems (as far
from equilibrium systems) normativity emerges in nature. Normative
asymmetry (adaptive/maladaptive, true/false, etc.) is transitive from
the asymmetry between energy-well stability (rocks, atoms, etc.) and
far-from-equilibrium stability. Functional
normativity is thus naturalized: it is the very system who
determines and specifies it, not an external observer attributing
functions to structures and imposing a normative criteria according
to its correspondence with states of affairs in the world nor
on the basis of the agents evolutionary history. Computational
and evolutionary functionalism provide, both,
heteronomous sources of normativity, unabling the very system
for error detection behaviour or any other kind of normative
re-organization.
If an autonomous system needs to recruit the same infrastructure to
achieve both constructive and interactive closure then the
space of possible biological organization becomes highly
constrained. This happens because metabolic reactions (constructive
processes) are slower than the reaction times required for available
interactive closure opportunities, specially those available for
fast body mouvements (motility) in big organisms (where the
relative difference in velocity between metabolic reactions and body
mouvement increases). Thus if a subset of the interactive closure is
achieved and controlled by a structure that instantiates processes
which are dynamically decoupled from the constructive ones, the space
of viable system organization is expanded. That's precisely the
origin of the nervous system: the new opportunities for survival
offered by the hierarchical decoupling of the nervous system, i.e.
behavioural control decoupled from metabolic (constructive)
constraints. Following (Moreno and Lasa, 2003) in this argument, the
relation between metabolic constructive processes (M) and the nervous
system (NS) is characterized by:
- Hierarchical decoupling of the NS from M: The NS is
hierarchically decoupled from M by the:
- Bottom-up, local, constructive causation of NS by M:
constructive processes produce a new dynamical domain, new
variables and relations between variables: the NS. The constructive
nature of this causation establishes the hierarchical aspect
of
the decoupling.
- Dynamic underdetermination of NS by M: the dynamic
state of the NS is underdetermined by metabolic dynamics
(decoupling).
- Downward causal dependency of M on NS: Because the NS
performs interactive functionality for the self-maintenance of the
system, M depends of the proper functioning of NS.
- Global and dynamic meta-regulation of NS by M: Although
dynamically underdetermined by M, because the NS's functionality is
defined by its interactive contribution to self-maintenance (and
this must ultimately be evaluated by M) M establishes the
metaestability condition for the NS. M does not directly evaluate
NS's dynamics but the interactive closure: i.e. the input of matter
and energy it gets from the environment.
We can now abstract a second domain in biological systems
(hierarchically decoupled from basic autonomy): the domain of
the organism's behavioural adaptive dynamics, specified by the
dynamical coupling between the embodied nervous system and the
environment and the metabolic meta-evaluation of that coupling.
This new dynamic domain, decoupled from local metabolic processes,
provides a qualitative lower level boundary for the characterization
of the specificity of cognition and allows for specific dynamical
modelling of adaptive behaviour. It is in this modelling that we will
be able to define behavioural adaptive functionality and thus a new
level of autonomy.
Dynamically considered metabolism only acts as a set of control
parameters for the nervous system; the behavioural domain is
dynamically blind to metabolism's constructive functioning. Thus
the constructive processes of basic autonomy can be modelled as a set
of essential variables which tend to stay away from equilibrium;
representing the cohesive limits of constructive processes and
their interactive closure conditions. A similar approach was already
taken by Ashby (1952) half a century ago (from whom we have taken
the term essential variables) and recently recovered by Beer (1997)
and Di Paolo (2003) in (evolutionary) simulation modelling of
adaptive behaviour. The dynamical autonomy of the behavioural domain
allows for a naturalistically justified assumption of dynamical
system theory (DST) as the proper conceptual framework to think about
autonomy and cognition in this domain. If we model
a) the agent's NS and the
environment as coupled dynamical systems (situatedness),
b) coupled through sensory and motor transfer functions
(embodiment), and
c) the metabolic processes as essential (far from
equilibrium) variables only controllable from the environment and
signalling the NS;
we get that functionality and autonomy can be redefined in the
behavioural domain (see figure 1).
Figure:
Kinetic graph of a dynamical modelling of adaptive behaviour.
Adapted from Ashby (1952): a closed sensorimotor loop
(controlled by the nervous system)
traverses the environment affecting metabolic
processes, which, in turn feeds-back to the nervous system
acting as a meta-regulatory mechanism. Embodiment is
modelled by sensory (S) and motor (M) surfaces.
|
In the behavioural domain thus considered, a new level of autonomy
can be described, hierarchically decoupled but interlocked with basic
autonomy: behavioural adaptive autonomy.
We can now, in dynamical terms, explicitly define behavioural
adaptive autonomy as:
homeostatic maintenance of essential variables under
viability constraints [adaptivity] through self-modulating
behavioural coupling with the environment [agency],
hierarchically decoupled from metabolic (constructive) processes
[domain specificity].
This definition highlights three main aspects of behavioural adaptive
autonomy:
- Adaptivity:
- Homeostatic maintenance of essential
variables under viability constraints assures a naturalized
and autonomous criteria for (adaptive) functionality. Next section
will further analyze the consecuences of functionality thus
considered.
- Agency:
- Self-modulation or self-restructuring of the
interactive coupling provides a criteria for autonomous
functionality (agency), excluding external contributions to
adaptation such as parents' care. Because the state of essential
variables is only accessible for the agent (through internal
sensors: level of glucose, feeling of hot, pain, etc.) the
homeostatic regulation must be guided by the agent's nervous system
and not by the environment. Thus the NS needs to evaluate it's
structural coupling through value signals from the essential
variables. This way a value system guided by the state of
essential variables and acting as metaestability condition for
structural plasticity of sensorimotor transformations becomes a
fundamental component of behavioural autonomy, and a defining
component of agency. The higher the
agent's capacity for adaptively guided self re-structuring
(plasticity) the higher it's behavioural adaptive autonomy and
hence its agency.
- Domain specificity:
- The hierachical decoupling of the
nervous system from metabolic processes provides a naturalized
criteria for the domain specificity of behavioural autonomy,
distinct form other adaptive domains in nature (bacterian
networks, plants, etc.). This domain specificity should not
be considered as independency but as hierarchical decoupling
(explained above), which allows for a justified specific modelling
of behavioural autonomy separated from local construtive
aspects. Two kinds of autonomy are interlocked
here: basic autonomy and behavioural autonomy. Both domains are
mutually required, the behavioural domain satisfies interactive
closure of basic autonomy and basic autonomy constructs the body
and neural variables defining the behavioural domain while acting
as a modulator of the structure of behavioural autonomy (see
figure 2).
Figure:
Basic and behavioural autonomy interlocked.
|
Functionality, in the behavioural domain thus considered, can be
defined as the mapping between agent-environment coupling and the
essential variables. Normativity is transitive from basic autonomy to
the behavioural domain through the maintenance of essential variables
under viability constraints. Thus normative functionality
(adaptation) is a mapping between agent-environment coupling
and the maintenance of essential variables under viability
constraints.
Because this definition of function does not compromise
any structural decomposition in functional primitives (unlike
traditional functionalism), a dynamical approach to behavioural
functionality can hold two kinds of emergence3:
- a) Structural emergence:
- If the agent's
structure is causally integrated (and the NS often is), i.e.
interactions between components are non-linear and components are
highly inter-connected, functional decomposition of components
(localisation) is not possible. The functionality of the system
emerges from local non-linear interactions between
components. FALTA EXPANDIR UN POCO CON LA DEF DE EMERGENCIA DE
BECHTEL
- b) Interactive emergence:
- Because
essential variables are non-controlled variables for the agent,
functionality is interactively emergent (Hendriks-Jansen, 1996; Steels, 1991), not in the trivial sense that essential variables
need external input, but in the sense that achieving this often
requires closed sensorimotor loops for the agent to enact the
necessary sensorimotor invariants to control essential
variables.4
What this double emergent condition shows is that the way the
specific adaptive function is achieved involves a dynamic
coupling between agent and environment where no particular
decomposable structure of the agent can be mapped into functional
components. This way holism is preserved as a characterizing
condition in complex systems but functionality can still be
explicitly defined. Localisation is not a valid explanatory
strategy but explanation is still possible (at least in principle).
And this is so because functionality is the outcome of a
process (not a structural relation between components): the
dynamical coupling between agent and environment.
herre
Now, the problem with behavioural adaptive autonomy is
the problem of a higher characterization and development of its
understanding, specially in relation to its self-regulating,
emergent and complex nature which does not allow for a
localisationist program to succeed: i.e. functional and structural
decomposition and aggregative causal abstraction of mutual relations
(Bechtel and Richardson, 1993).
When localisation is thrown away the locus of
philosophical enquiry regarding the nature and origin of cognition
(if a naturalist and biologically inspired philosophical approach is
to be adopted) is displaced towards;
- the specification of the dynamic organization of lower
level mechanisms capable of implementing behavioural adaptive
autonomy and FALTA EXTENDER
- the search for the nature of intermediate explanatory patterns
between the agent-environment structural coupling and the maintenance
of essential variables under viability constraints: traditional
concepts (information, representation, memory, processing, etc.)
should be dynamically grounded.
This task is genuinely philosophical and distinct from specific
modelling of biologicall targets.
A-life (Moreno, 2000; Langton, 1996; Dennet, 1995) and, more specifically,
evolutionary simulation modelling5 becomes a
mayor philosophical tool here, not for a mere synthesising of
behavioural autonomy but for philosophical research through opaque
though experiments (Di Paolo et al., 2000) with conceptually
(dynamically) complex systems, produced, implemented and manipulated
in a computer. The simulation acts as an artefactual blending
(Fauconnier and Turner, 1998) between lower level neural mechanistic
concepts and global functional conceptualization of behaviour
(Barandiaran and Feltrero, 2003).
Evolutionary simulation modelling works by:
- Definition of a set of body, environment and neural structures
(unspecified on their parameter values). Neural structures are
abstractions of a set of lower level neural mechanisms from
neuroscientific models (functionally unspecific),
and body structures are a set of robotic idealizations.
- Artificial evolution of parameters according to a given
fitness function.
- Reproduction/simulation of system behaviour with numerical
methods allowing for qualitative analysis of complex dynamical
systems.
Highly connected CTRNNs (continuous time recurrent neural networks)
are used in evolutionary robotics to model de agent's control
architecture. The dynamics of such networks are highly complex,
capable (in principle) to emulate any other dynamical system with a
finite number of variables (Funakashi and Nakamura, 1993).
Because the lower level mechanisms are functionally unspecific and
artificial evolution is used to achieve emergent functionality,
evolutionary simulation modelling has long being used as tool to
produce proofs of concept regarding the relation between lower level
mechanisms and global behaviour. Examples of such proofs of concept
include the production of minimally cognitive behaviour without
explicit internal representations (Beer, 2001), autonomous learning
in neural networks without synaptic plasticity (Tuci et al., 2002) or
the achievement of functional readaptation to sensorimotor disruption
(through homeostatic synaptic plasticity) without disruption ever
being present on the evolutionary history of the simulated agent
(Di Paolo, 2000). This simulation models do not pretend to model any
specific biological target, but are rather used as philosophical or
intratheoretical experiments and their consequences for the
philosophy of biology and philosophy of the mind are significant on
that they keep testing theoretical assumptions and illustrating
conceptual re-organization.
In addition to this synthetic bottom-up methodology other
analytic tools should be philosophically tuned. Complexity measures
to understand functional integration in neural processes
(Tononi et al., 1998) are producing interesting results. An early
exploratory example of such methodology is provided by
Seth (2002), fusioning both evolutionary simulation modelling
and complexity measures of neural network dynamics. Complexity
measures showed that when the evolved networks dynamics are analysed
by random activation of neurons results are significantly different
than analysed when coupled with the environments for which they where
evolved. The experiment aiming to demonstrait that, at least in one
case, complex behaviour requires complex mechanisms, ends up showing
that structural analysis of network connectivity is never enough and
that coupling (situatedness) becomes, once again, a fundamental
characteristic of behaviour. FALTA ELABORAR UN POCO MÁS EL
TRATAMIENTO DE LA INTEGRACIÓN EN EDELMAN.
The strength of the proposed dynamical perspective on behavioural
adaptive autonomy (as a lower boundary condition for a naturalist
characterization of cognition) is given by the shift from:
- viewing cognition as computations between
`representational' automaton states, whose representational
normativity is fixed by an heteronomously interpreted
functional equivalence with states of affairs in the
world
to:
- and interactive dynamical process whose normativity
is given by its satisfaction of closure criteria and
functionality is grounded on the embodied and situated
nature of behavioural dynamics (structurally and interactively
emergent and capable of self-restructuring according to the
metabolic evaluation of the interactive coupling).
But further discussion and clarification is required in
this direction. Although behavioural adaptive autonomy satisfies the
goals mentioned in section 2, further developments in, at least, two
directions seem plausible.
Adaptive behavioural autonomy underdetermines cognitive
behaviour. Homeostatic maintenance of essential variables and self
re-structuring capacity is necessary but not sufficient for a
characterization of a gradual notion of cognition. In this sense we
believe that the work of Christensen and Hooker (2002) on
self-diretedness is a natural step forward. FALTA EXTENDER ESTO UN
POCO Y VOLVER A HABLAR DE INFORMACIÓN BREVEMENTE, INCLUYENDO UNA
MENCIÓN A "INFORMATION AND AUTONOMY" CON ALVARO.
Could a characterization of cognition come from other sources rather
than maintenance of essential variables within viability
constraints? Could a sort of sensorimotor coherence become an
alternative source of normativity? possibly a kind of minimal
structural metaestability condition for any kind of coherent
behaviour (whether this behaviour is adaptive or not)?
If something different to behavioural adaptivity was necessary for
complex cognitive behaviour to happen (e.g. the metaestability above
mentioned), that condition would enable adaptive behaviour, but not
everything enabled by that condition would be adaptive. Because that
condition would be necessary for cognition it could be considered
normative (as condition of possibility) and thus a new normative
domain would appear decoupled from interactive closure criteria. This
will admit non adaptive behaviour to still be cognitive.
An interesting line of research has recently been proposed by
Di Paolo (2003) in this direction. Di Paolo argues that behaviour
itself is underdetermined by survival conditions and proposes
habit formation as the origine of intentionality. Habits are
self sustaining dynamic structures of behavioural patterns,
sensorimotor invariants homeostatically maintained by neural
organization. Homeostatically controlled synaptic plasticity
(Turrigliano, 1999) could be a relevant neural organization leading
to such autonomy of behavioural patterns; as demonstrated by
Di Paolo (2000).
Rather
than providing conclusive results, what such research on synthetic
bottom-up simulation modelling is showing (within a dynamical
approach to cognition) is that the time is ready to address
important philosophical issues in a workable methodological and
conceptual framework for the study of behavioural autonomy. By
providing an explicit definition of behavioural adaptive autonomy in
this framework we hope to have contributed something in this
direction.
-
Ashby, W. (1952).
- Design for a Brain. The origin of adaptive behaviour.
Chapman and Hall, 1978 edition.
-
Barandiaran, X. and Feltrero, R. (2003).
- Conceptual and methodological blending in cognitive science. The
role of simulated and robotic models in scientific explanation.
Accepted paper for the 12th International Congress of Logic,
Methodology and Philosophy of Science, Oviedo (Spain), August 7-13, 2003.
-
Bechtel, W. and Richardson, R. (1993).
- Discovering Complexity. Decomposition and Localization as
strategies in scientific research.
Princeton University Press.
-
Beer, R. D. (1996).
- Toward the evolution of dynamical neural networks for minimally
cognitive behaviour.
In Maes, P., Mataric, M., Meyer, J. A., Pollack, J., and Wilson, S.,
editors, From Animals to Animats 4: Proceedings of the Fourth
International Conference on Simulation of Adaptive Behaviour, pages
421-429. Harvard, MA: MIT Press.
-
Beer, R. D. (1997).
- The Dynamics of Adaptive Behavior: A research program.
Robotics and Autonomous Systems, 20:257-289.
-
Beer, R. D. (2001).
- The dynamics of active categorical perception in an evolved model
agent.
submitted to Behavioral and Brain Sciences.
Downloaded on 13/3/02 from http://vorlon.cwru.edu/~beer/.
-
Bickhard, M. H. (2000).
- Autonomy, Function, and Representation.
Communication and Cognition - Artificial Intelligence,
17(3-4):111-131.
Special Issue on: The contribution of artificial life and the
sciences of complexity to the understanding of autonomous systems. Guest
Editors: Arantza Exteberria, Alvaro Moreno, Jon Umerez.
-
Block, N. (1996).
- What is Functionalism. Online revised entry on functionalism in the
.
In Borchert, D., editor, The Encyclopedia of Philosophy
Supplement. MacMillan.
URL:
http://www.nyu.edu/gsas/dept/philo/faculty/block.
-
Christensen, W. and Bickhard, M. (2002).
- The process dynamics of normative function.
Monist, 85 (1):3-28.
-
Christensen, W. and Hooker, C. (2002).
- Self-directed agents.
Contemporary Naturalist Theories of Evolution and
Intentionality, Canadian Journal of Philosophy, 31 (special issue).
-
Clancey, W. (1989).
- The Frame of Reference Problem in Cognitive Modeling.
In Arbor, A., editor, Proceedings of the 11th Annual Conference
of The Cognitve Science Society, pages 107-114. Lawrence Erlbaum
Associates.
-
Collier, J. (1999).
- Autonomy and Process Closure as the Basis for Functionality.
In Chandler, J.L.R./van de Vijver, G., editor, Closure:
Emergent Organizations and their Dynamics. Volume 901 of the New York Academy
of Sciences.
-
Cristensen, W. and Hooker, C. (1999).
- An Interactivist-Constructivist Approach to Naturalism,
Intentionality and Mind.
In Presented to Naturalism, Evolution, and Mind. The 1999 Royal
Institute of Philosophy Conference. University of Edimburg.
-
Dennet, D. (1995).
- Artificial Life as Philosophy.
In Langton, C., editor, Artificial Life. An overview, pages
291-2. MIT, Cambridge, MA.
-
Di Paolo, E. (2000).
- Homeostatic adaptation to inversion of the visual field and other
sensorimotor disruptions.
In Meyer, J.-A., Berthoz, A., Floreano, D., Roitblat, H., and Wilson,
S., editors, From Animals to Animats 6: Proceedings of the Sixth
International Conference on Simulation of Adaptive Behavior, pages
440-449. Harvard, MA: MIT Press.
-
Di Paolo, E. (2003).
- Organismically-inspired robotics: homeostatic adaptation and
teleology beyond the closed sensorimotor loop.
Working paper.
-
Di Paolo, E., Noble, J., and Bullock, S. (2000).
- Simulation Models as Opaque Thought Experiments.
In Bedau, M., McCaskill, J., Packard, N., and Rasmussen, S., editors,
Artificial Life VII: The 7th International Conference on the Simulaiton
and Synthesis of of Living Systems. Reed College, Oregon, USA.
-
Fauconnier, G. and Turner, M. (1998).
- Conceptual Integration Networks.
Cognitive Science, 22(2):133-187.
-
Funakashi, K. and Nakamura, Y. (1993).
- Approximation of dynamical systems by continuous time recurrent
neural networks.
Neural Networks, 6:1-64.
-
Gilbert, S. F. and Sarkar, S. (2000).
- Embracing Complexity: Organicism for the 21st Century.
Developmental dynamics, 219:1-9.
-
Harvey, I., Husbands, P., Cliff, D., Thompson, A., and Jakobi, N. (1997).
- Evolutionary Robotics: the Sussex Approach.
Robotics and Autonomous Systems, 20:205-224.
-
Hendriks-Jansen, H. (1996).
- In praise of interactive emergence, or why explanations don't have
to wait for implementations.
In Boden, M., editor, The Philosophy of Artificial Life,
pages 282-299. Oxford University Press, Oxford.
-
Langton, C. (1996).
- Artificial Life.
In Boden, M., editor, The Philosophy of Artificial Life,
pages 39-94. Oxford University Press, Oxford.
-
Maes, P., editor (1991).
- Designing Autonomous Agents.
MIT Press.
-
Maturana, H. and Varela, F. (1980).
- Autopoiesis. The realization of the living.
In Maturana, H. and Varela, F., editors, Autopoiesis and
Cognition. The realization of the living, pages 73-138. D. Reidel
Publishing Company, Dordrecht, Holland.
-
Millikan, R. G. (1989a).
- Biosemantics.
Journal of Philosophy, 86 Issue 6 (June):281-297.
-
Millikan, R. G. (1989b).
- In defense of proper functions.
Philosophy of Science, 56:288-302.
-
Moreno, A. (2000).
- Artificial Life as a bridge between Science and Philosophy.
In Bedau, M.A., M. J. P. N. and Rasmussen, S., editors, Artificial Life VII: The 7th International Conference on the Simulaiton and
Synthesis of of Living Systems. MIT Press.
-
Moreno, A. and Lasa, A. (2003).
- From Basic Adaptivity to Early Mind.
Evolution and Cognition, 9(1):in press.
-
Nolfi, S. and Floreano, D. (2000).
- Evolutionary Robotics: The Biology, Intelligence and Technology
of Self-Organizing Machines.
MIT Press.
-
Ruiz-Mirazo, K. and Moreno, A. (2000).
- Searching for the Roots of Autonomy: the natural and artificial
paradigms revisited.
Artificial Intelligence, 17 (3-4) Special issue:209-228.
-
Seth, A. K. (2002).
- Using dynamical graph theory to relate behavioral and mechanistic
complexity in evolved neural networks.
Unpublished. Url:
http://www.nsi.edu/users/seth/Papers/nips2002.pdf.
-
Slocum, A. C., Downey, D. C., and Beer, R. D. (2000).
- Further experiments in the evolution of minimally cognitive
behavior: From perceiving affordances to selective attention.
In Meyer, J., Berthoz, A., Floreano, D., Roitblat, H., and Wilson,
S., editors, From Animals to Animats 6: Proceedings of the Fourth
International Conference on Simulation of Adaptive Behavior, pages
430-439. Harvard, MA: MIT Press.
-
Steels, L. (1991).
- Towards a Theory of Emergent Functionality.
In Meyer, J. and Wilson, R., editors, Simulation of Adaptive
Behaviour, pages 451-461. MIT Press.
-
Tononi, G., Edelman, G., and Sporns, O. (1998).
- Complexity and coherency: integrating information in the brain.
Behavioural and Brain Sciences, 2(12):474-484.
-
Tuci, E., Harvey, I., and Quinn, M. (2002).
- Evolving integrated controllers for autonomous learning robots using
dynamic neural networks.
In Proceedings of The Seventh International Conference on the
Simulation of Adaptive Behaviour (SAB'02).
-
Turrigliano, G. (1999).
- Homeostatic plasticity in neuronal networks: The more things change,
the more they stay the same.
Trends in Neuroscience, 22:221-227.
-
van Gelder, T. (1998).
- The dynamical hypothesis in cognitive science.
Behavioural and Brain Sciences, 21:615-665.
-
Varela, F. (1979).
- Principles of Biologicall Autonomy.
North-Holland, New York.
- Environment (1):
- The environment take in isolation will be
defined by
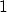 ; where
; where
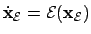 is the state vector of the environment
and
is the state vector of the environment
and 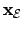 the function gouverning its change. If the agent
can induce changes in the environment we can open the model and in
the following way:
the function gouverning its change. If the agent
can induce changes in the environment we can open the model and in
the following way:
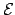 ; where
; where
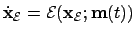 is the agent's motor activity vector acting on the
environment.
is the agent's motor activity vector acting on the
environment.
- Agent (1):
- We take the agent to be the mechanism specifying
agency in the adaptive system (i.e. not considering metabolic
processes, constructive autonomy). Thus:
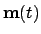 ; where
; where
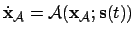 is the state vector of the agent,
is the state vector of the agent,
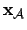 the lows governing the agents variables and
the lows governing the agents variables and
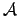 the sensory input vector to the agent.
the sensory input vector to the agent.
- Essential variables:
- Our modelling only captures relevant
operational structures for adaptive behaviour. In this
context we model essential variables as enclosing mainly
thermodynamic conditions in relation to the agent as a
far from equilibrium system. Essential variables must be under
viability constraints for the system to maintain its biological
organization. Essential variables are non controled variables; i.e.
the agent can only modulate those variables through the environment.
Thus we define the essential variable vector as
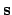 where the
input to essential variables
where the
input to essential variables
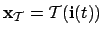 .
Where
.
Where 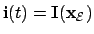 is a function of the environment determining the
input to essential variables. We shall, thus, define the rate of
change of essential
variables as
is a function of the environment determining the
input to essential variables. We shall, thus, define the rate of
change of essential
variables as
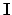 , where
, where
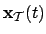 .
.
- Agent-Environment coupling:
- Because the sensory input is a
function of the environment (in relation to the agent) we can
specify that
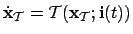 .
And converselly:
.
And converselly:
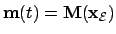 .
.
- Environment (2):
- Thus we can re-define the environment
(introducing the agent's effect) as
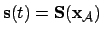 .
.
- Agent (2):
- If we consider that the agent
might get some signal vector
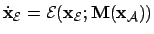 , so that
, so that
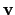 from its essential variables
and considering the agent-environment coupling we get
from its essential variables
and considering the agent-environment coupling we get
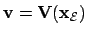 .
.
- Adaptive System:
- We can define the adaptive system as the
coupling between
and
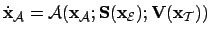 .
.
- Agent-Environment coupling (2):
- The agent-environment
coupled system will be:
- Universe:
-
- Sensorimotor transformation:
- Sensorimotor transformations
are the trajectories of
- Behaviour:
- Metalevel description of agent-environment
interaction. Behaviours must be defined in the
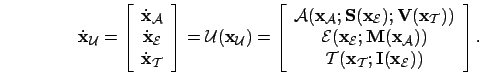 domain,
adaptive behaviour in the
domain,
adaptive behaviour in the 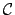 domain.
domain.
- Organization:
- The set of metainvariant relations between the
variables defining a system: the functions governing variables.
- Structure:
- The set of invariant relations between the
variables defining a system: it is composed of functions and
parameters. When some variables of a system remain stable for long
periods of time (comparing to the rate of change of other variables)
they can be considered parameters; lets call them weak
parameters.
- Plasticity:
- Capacity of a given organization to induce
structural changes, i.e. weak parameter changes.
- Adaptive (normative) Function:
- Because de Agent is a far
from equilibrium system it must keep some of its essential variables
under viability constraints activelly (since
they will tend to decay according to the second
law of thermodynamics). Some essential variables represent (in
the model) this thermodynamic condition, from which the agent gets
indications trough
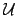 . Other essential variables (like body
integrity) must be keept within viability constraints during the
interaction processes. Thus adaptive functionality
. Other essential variables (like body
integrity) must be keept within viability constraints during the
interaction processes. Thus adaptive functionality 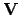 can
be defined as a mapping from the coupled agent-environment system to
the essential variables, so that the essential variables are keep
between viability constraints; i.e.
can
be defined as a mapping from the coupled agent-environment system to
the essential variables, so that the essential variables are keep
between viability constraints; i.e.
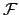 ; where
; where
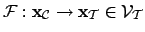 is the
viability subspace of the essential variables. FALTA: SOME
ESSENTIAL VARIABLES DON'T DECAY (BODY INTEGRITY).
TAMBIÉN HAY QUE DISTINGUIR ENTRE FUNCIÓN DE UN PROCESO Y FUNCIÓN
PROPIA DE UNA ESTRUCTURA
is the
viability subspace of the essential variables. FALTA: SOME
ESSENTIAL VARIABLES DON'T DECAY (BODY INTEGRITY).
TAMBIÉN HAY QUE DISTINGUIR ENTRE FUNCIÓN DE UN PROCESO Y FUNCIÓN
PROPIA DE UNA ESTRUCTURA
- Behavioural Adaptive Autonomy:
- An adaptive system is
autonomous if it is capable of homeostatically maintaining
under viability constraints throught
self-modulating behavioural coupling with the environment. Adaptive
functionality is always the outcome of the agent-environment
coupling, thus adaptive autonomy is a continous measure of the
agent's active contribution to the satisfaction of adaptive
functionality.
- Mechanistic Explanation:
- The answer to the question ``which
subset of is causally relevant to the performance of a
function?''. FALTA INTRODUCE HYPERDESCRIPTIONS HERE (READ CHRIS).
- Localization:
- Functional components' mapping into structural
components. Localization requires structure and functionality to be
decomposed, which might not be always possible if struture is
integrated (i.e. if interaction between components is higher than
within components).
Footnotes
- ...
normativity1
- Contributions of authors to the development of
the concept of autonomy (in relation to autopoiesis, cognition,
functionality, normativity, etc.) did not happen in isolation,
crossreferencing and collaboration has been a common practice
so that specific contributions as outlined above shouldn't be taken
too rigorously.
- ...
laws2
- Although
new physical laws will never be created by an organism, or any other
system, it can always generate new constraints and internal control
mechanisms
- ... emergence3
- We are here
talking of weak emergence in the sense of an holistic, recursive and
distributed causal structure.
- ...
variables.4
- Very often interactive emergence reinforces
structural emergence because ``interactions between separate
sub-systems are not limited to directly visible connecting links
between them, but also include interactions mediated via the
environment'' (Harvey et al., 1997, p.205)
- ... modelling5
- Evolutionary robotics
(Nolfi and Floreano, 2000; Harvey et al., 1997) and Randall Beer's
minimally cognitive behaviour program (Beer, 2001,1996; Slocum et al., 2000) being the major exponents here.
xkrop
2003-11-18
![\begin{displaymath}
\dot{\mathbf{x}}_{\mathcal{U}} =
\left[\begin{array}{c}
\...
...};
\mathbf{I}(\mathbf{x}_{\mathcal{E}})) \end{array} \right].
\end{displaymath}](img23.png)